|
|
| (15 intermediate revisions by the same user not shown) |
| Line 1: |
Line 1: |
| | {{unfinished}} | | {{unfinished}} |
| | ==Central Nervous System== | | ==Central Nervous System== |
| − | ===Brain=== | + | ===[[Equine Brain - Horse Anatomy|Brain]]=== |
| − | The brain is responsible for co-ordinating, integrating and controlling the rest of the nervous system. The brain is divided into several parts. Based on phylogenetic development, it can be divided into the forebrain, midbrain and hindbrain. Based on gross anatomy, it can be divided into the cerebrum, cerebellum and brainstem. The brain is enclosed within the cranial cavity of the skull.
| |
| − | ====Forebrain====
| |
| − | The ''forebrain (proencephalon)'' is the largest part of the brain, most of which is ''cerebrum''. Other important structures found in the forebrain include the ''thalamus'' , the ''hypothalamus'' and the ''limbic system''. The cerebrum is divided into two cerebral hemispheres connected by a mass of white matter known as the ''corpus callosum''. Each hemisphere is split into four lobes; the ''frontal'', ''parietal'', ''occipital'' and ''temporal'' lobes. The surface of each hemisphere is made up of grey matter known as the ''cerebral cortex'' and is folded to increase the surface area available within the skull. The cortex has roles within perception, memory and all higher thought processes. Inside the cortex is the ''white matter'', within which are a number of nuclei (grey matter), known as the ''basal nuclei''. The basal nuclei receive information from the cortex to regulate skeletal movement and other higher motor functions.
| |
| | | | |
| − | The thalamus functions to relay sensory information to the cerebral cortex and the hypothalamus, regulating visceral functions including temperature, reproductive functions, eating, sleeping and the display of emotion. The limbic system describes a collection of structures within the forebrain, including the ''amygdala'' and '' hippocampus'', also known as the 'emotional brain'. It is important in the formation of memories and in making decisions and learning.
| + | ===[[Equine Cranial Nerves - Horse Anatomy|Cranial Nerves]]=== |
| − | =====Thalamus===== | |
| − | The thalamus is comprised of two oval shaped structures, connected by the '''interthalamic adhesion'''.
| |
| − | The thalamus has many functions. It is involved in processing and relaying sensory information selectively to various parts of the cerebral cortex. All senses from lower centres, including auditory, somatic, visceral, gustatory and visual systems, have input to the thalamus before being relayed to the appropriate areas within the cerebral cortex. The thalamus plays a major role in regulating arousal, levels of consciousness and levels of activity.
| |
| | | | |
| − | =====Hypothalamus===== | + | ===[[Vasculature of the Equine Brain - Horse Anatomy|Vasculature of the Brain]]=== |
| − | The hypothalamus is located on the floor of the diencephalon, beneath the thalamus. It communicates with the cerebral cortex, thalamus, [[Equine Endocrine System - Horse Anatomy#Pituitary Gland|pituitary gland]] and other areas of the brain via the '''infundibular stalk'''.
| |
| | | | |
| − | The function of the hypothalamus is mainly related to the overall regulation of the [[Equine Endocrine System - Horse Anatomy|Endocrine System]]. It also controls and integrates the autonomic nervous system, therefore controlling heart rate, blood pressure, body temperature and gastrointestinal secretions. The hypothalamus is closely related to the [[Equine Endocrine System - Horse Anatomy#Pituitary Gland|pituitary gland]], controlling a large proportion of the activity going to it. For a more detailed analysis of the function of this part of the brain, please use the link: [[Hypothalamus - Anatomy & Physiology|Hypothalamus Anatomy and Physiology]].
| + | ===[[Equine Spinal Cord - Horse Anatomy|Spinal Cord]]=== |
| | | | |
| − | =====Pituitary===== | + | ===[[Equine Meninges - Horse Anatomy|Meninges]]=== |
| − | The function of the pituitary is mainly related to the production of hormones as part of the Endocrine System. For further information on the pituitary gland please use this link: [[Equine Endocrine System - Horse Anatomy#Pituitary Gland|Equine Pituitary Gland]].
| |
| | | | |
| − | =====Cerebral Cortex===== | + | ===[[Equine Cerebrospinal Fluid - Horse Anatomy|Cerebrospinal Fluid]]=== |
| − | This cerebrum is the largest part of the brain, divided by the '''median fissure''' into two cerebral hemispheres. The cerebral hemispheres are connected by the '''corpus callosum'''. The outer layers of the cerebrum are made up of grey matter. Grey matter is formed by neurons and their unmyelinated fibres. The white matter below the grey matter of the cortex is formed predominantly by myelinated axons (myelin is white in appearance). The surface of the cerebral cortex is folded, the numerous folds are known as '''gyri''' which greatly increase the surface area. Grooves between these gyri are known as '''sulci'''; more than two thirds of the surface is within sulci. The cerebral cortex is connected to structures such as the thalamus and the basal ganglia, sending information to them along ''efferent'' connections and receiving information from them via ''afferent'' connections. Most sensory information is routed to the cerebral cortex via the thalamus. The cortex is commonly described as comprising three parts; sensory, motor and association areas.
| |
| | | | |
| − | The '''sensory areas''' receive and process information from the senses. Inputs from the thalamus are called ''primary sensory areas''. Vision, hearing, and touch are processed by the primary visual cortex, primary auditory cortex and primary somatosensory cortex respectively. The two hemispheres of the cerebral cortex receive information from the opposite (contralateral) side of the body. There are a number of anatomical areas of the brain responsible for organising this sensory information. The ''parietal lobe'' is located within the dorsocaudal aspect of the cortex. The ''temporal lobes'' are located laterally and the ''occipital lobes'' are located in the caudal most aspect of the cortex. The ''frontal lobe'' or ''prefrontal association complex'' is involved in planning actions and movement.
| + | ===Clinical Links=== |
| | + | *[[Equine Protozoal Myeloencephalitis]] |
| | + | *[[Equine Herpesvirus 1]] |
| | + | *[[Polyneuritis Equi]] |
| | | | |
| − | The '''motor cortex''' areas of the brain are located in both hemispheres of the cortex and are shaped like a pair of headphones stretching from ear to ear. The motor areas are related to controlling voluntary movements, especially fine movements. There are two main types of connection between the motor cortex and motor neurones found in the ventral horn of the spinal cord; the ''Pyramidal tracts'' and the ''Extrapyramidal tracts''. | + | ==Peripheral Nervous System== |
| − | *'''Pyramidal tract''' connections are direct with no synapses in the brain stem. Axons pass through the ventral aspect of the medulla oblongata. The pyramidal tracts are responsible for aspects of fine motor skills that require a degree of conscious thought and concentration.
| + | ==Introduction== |
| − | *The '''extrapyramidal tracts''' pass through the medulla oblongata outside the ventral pyramidal tracts and have synapses within the brain stem nuclei. These synapses make it possible for signals travelling down the extrapyramidal horns to be influenced by other areas of the brain including the cerebrum. The extrapyramidal tracts are generally responsible for activation of larger muscle groups and often work in a coordinated manner to achieve smooth synchronous movements.
| + | The '''Peripheral Nervous System''' is made up of cranial and spinal nerves. Spinal nerves are named after the vertebra immediately above it, except for '''cervical vertebra'''. There are '''7''' cervical vertebrae and '''8''' cervical spinal nerves. The peripheral nervous system can be divided into the '''somatic nervous system''' and '''autonomic nervous system'''. The somatic nervous system co-ordinates body movements and also receives external stimuli. It regulates activities that are under conscious control. The autonomic nervous system subdivided into the '''sympathetic nervous system''', '''parasympathetic nervous system''', and enteric division. The sympathetic nervous system is the '''‘fight or flight’''' system which comes into role when an animal is under threat, its main neurotransmitter is '''adrenaline'''. The parasympathetic nervous system is the '''‘rest and digest’''' system which is responsible for digestion. Its main neurotransmitter is '''acetylcholine'''. |
| − | | + | ==Structure== |
| − | =====Limbic System=====
| + | [[Image:WIKIVETperipheralnervestructure.jpg|thumb|right|150px|Peripheral Nerve Structure. Sophie Stenner, RVC 2008]] |
| − | The Limbic system is made up of parts of the brain bordering the '''corpus collosum'''. The Limbic system contains areas of '''cerebral cortex''', the '''cingulate gyrus''' (dorsally), the '''parahippocampus gyrus''' (ventrally), the '''amygdala''', parts of the '''hypothalamus''' (mamillary body) and the '''hippocampus'''. The limbic system is principally responsible for emotions and the various types of emotion can affect the activity of the ''[[Autonomic_Nervous_System_-_Anatomy_%26_Physiology|Autonomic Nervous System]]'', facilitated by the hypothalamus. | + | Nerve fibres reside in a connective tissue matrix called the '''endoneurium''' and are gathered together into bundles or fascicles defined by a second connective tissue layer called the '''perineurium'''. Groups of fascicles are then gathered together in a third connective tissue layer called the '''epineurium'''. Thus, peripheral nerves have a '''three-tiered hierarchical arrangement of connective tissue'''. '''Renaut bodies''' are loose, cigar-shaped whorls of extracellular matrix within fascicles that are common in equine nerves at points of stress or compression. |
| − | | + | ==Nerve Fibre== |
| − | =====Olfactory Bulb=====
| + | The nerve fibre consists of the impulse-carrying axon, which is surrounded by an ensheathing cell, the [[#The Schwann cell|Schwann cell]], which in turn is surrounded by an acellular basal lamina that is continuous along the length of the nerve. Nerve fibres come in various discrete diameter groups, which are reflected in their conduction velocities. The larger the diameter the more rapid the rate of impulse conduction. Particular targets or receptors are associated with axons of a particular diameter. Axons connected to muscles spindles have a large diameter (20 µm) and conduct at 120 m/s whilst the smallest myelinated fibres are about 1µm and conduct at around 6 m/s. The smallest fibres of all are the unmyelinated fibres (the high-threshold sensory afferents, or C-fibres, and post-ganglionic autonomies) and have a diameter of between 1 and 0.1 µm. These fibres do not conduct by saltatory conduction and have very slow conduction rates of around 0.5 m/s. |
| − | The olfactory bulb is responsible for olfaction and the bulb itself is located within the rostral forebrain area, supported by the ''cribiform plate'' and the ''ethmoid bone''. The olfactory nerves are connected directly to the limbic system.As a result, olfaction plays a central role and is particularly important in regulating/stimulating sexual behaviour.
| + | ===Axon=== |
| − | | + | Axons have an outer membrane called the '''axolemma''' and within this there is the '''axoplasm''' which is continuous with the cytoplasm of the [[Neurons - Anatomy & Physiology|neuron]]. There are no ribosomes, either free or attached to endoplasmic reticulum in axons and therefore, no protein synthesis. Protein synthesis takes place within the cell body and some dendrites and all protein replacement required for the maintenance of the axon depends on proteins being imported from the cell body. A critical feature of the axon is its '''cytoskeleton''', which consists of two key elements; '''neurofilaments''' and '''microtubules'''. '''Neurofilaments''' are intermediate filaments of about 10 nm diameter, and belong to the same class as other cytoskeletal proteins such as keratin, desmin, vimentin, or GFAP of astrocytes. Neurofilaments are formed from a triplet of polypeptide subunits of heavy (~ 200 kD), medium (~ 150 kD) and low (~ 60 kD) molecular weights. Typically, these subunits are heavily phosphorylated and are more numerous than microtubules, especially in large diameter axons, having a pivotal role in determining axon diameter. They are formed in the cell body, transported down the axon by axoplasmic transport and degraded in the terminals by Ca<sup>2+</sup> activated proteases. In other words, there is a constant turnover of neurofilament within the healthy axon. '''Microtubules''' within axons are similar to microtubules elsewhere, consisting of polymerised dimers of alpha and beta tubulin arranged as a hollow tube of about 28 nm. They are relatively abundant in smaller diameter axons, and are also synthesised in the cell body. An important component of the cytoskeleton are the '''microtubule associated proteins''' or MAP's and the tau proteins. These proteins are important in microtubule assembly and stability. Different classes of MAP's occur in the dendrites and the axons, and to some extent account for the different ultrastructural features that distinguish these two types of neuronal process. They form cross links between adjacent microtubules but also connect to neurofilaments and actin microfilaments, implying complex interactions between the various components of the axon skeleton. |
| − | ====Midbrain==== | + | ===Schwann Cell=== |
| − | The midbrain or mesencephalon represents the connection between the [[Hindbrain - Anatomy & Physiology#Hind Brain Structures & Functions|brain stem]] and the higher centres of the brain and is involved in most body systems including '''sleep/consciousness, vision, hearing and temperature regulation'''. | + | Myelination in the PNS is achieved by the '''Schwann cell''', a derivative of neural crest cells, which bud off from the neuroepithelium at a very early stage of neurogenesis. During development, Schwann cells engage many small axons and as axonal diameter increases, Schwann cells eventually relate with only a single axon c.f [[Neurons - Anatomy & Physiology#Oligodendrocytes|oligodendrocytes]]. This single axon is enveloped in a trough by the Schwann cell processes that engulf it and as the processes come together, an inner '''mesaxon''' is formed. The leading-edge process continues to move over the axon forming a spiral. Myelination, an extremely complex molecular process, occurs when the cytoplasm within the process is extruded allowing the internal surfaces of the membrane to come together as the '''major dense line''', the outer membrane apposition constituting the intraperiod line. The alternating pattern of these two form the lamellae of compacted myelin. The myelin sheath is attached to, and is an integral part of the Schwann cell on which it is dependent for its maintenance. |
| − | | |
| − | The midbrain is located between the diencephalon and the hind brain, or brain stem. More specifically, it can be found ventral to the [[Cerebrum|cerebral cortex]] and between the cerebral pedicles of the diencephalon and the pons. It is a relatively short portion of the upper brain stem and connects higher brain centres with the lower centres and spinal cord. | |
| − | | |
| − | The midbrain has a stratified structure comprising various layers including the '''tectum, tegmentum '''and '''cerebral peduncle'''. These structures are found in a dorsoventral sequence. The tectum lies dorsally to the cerebral aqueduct and it has four major rounded surface swellings; colliculi (see below). The tegmentum is the core of the midbrain and a large proportion of it is made up by the [[Sensory Pathways - Anatomy & Physiology#Ascending Reticular Formation (Spinoreticular Tract)|reticular formation]].
| |
| − | =====Cranial Nerve Nuclei=====
| |
| − | The major cranial nerve nuclei within the midbrain are the mesencephalic nuclei of the [[Cranial Nerves - Anatomy & Physiology|trigeminal nerves (V)]], the trochlear nuclei (IV), the principle and parasympathetic oculomotor nuclei [[Cranial Nerves - Anatomy & Physiology|(Cranial Nerve III)]], the 'red nuclei' (so named due to its pronounced vascularity) and the periaqueductal grey nuclei. The periaqueductal grey nucleus is a core of grey nervous tissue located adjacent to the cerebral aqueduct. The oculomotor nerve emerges from the mid brain rostral to the pons.
| |
| − | =====Cerebral Aqueduct=====
| |
| − | Within the lumen of the midbrain lies the cerebral aqueduct, which acts as a simple passage between the spinal cord and the third and fourth ventricles.
| |
| − | =====Colliculi=====
| |
| − | The tectum (roof) has four colliculi, two rostral and two caudal.
| |
| − | | |
| − | ======Caudal Colliculi======
| |
| − | The caudal colliculi are widely spaced and are joined by a substantial commissure. The caudal colliculi act as '''integration centres for auditory pathways'''. The caudal colliculi also have a further connection to the thalamus via the ipsilateral '''medial geniculate body'''. This body is effectively a swelling of the [[thalamus]].
| |
| − | | |
| − | ======Rostral Colliculi======
| |
| − | The rostral colliculi are placed closer together in comparison to the caudal. The rostral colliculi are also joined to the thalamus, but by the '''lateral geniculate bodies''' rather than the medial. The rostral colliculi help to '''integrate the visual pathways''' and also are involved in '''somatic reflexes''' that are caused by visual cues. The rostral colliculi have also been suggested to be involved in spatial integration.
| |
| − | | |
| − | =====Substantia Nigra===== | |
| − | The substantia nigra is a prominent area of the mid brain and is identifiable on cross sections by its darker pigmentation. This pigmentation is due to the gradual accumulation of pigmentation of neurons and is associated with basal nuclei within the tissue. The '''substantia nigra''' nuclei are involved in the '''control of voluntary movement'''.
| |
| − | | |
| − | =====Crura Cerebri=====
| |
| − | These are visible on the ventral surface of the mid brain and consist of fibre tracts that are in passage between the telencephalon and the brain stem. The oculomotor nerves (see above) also emerge in this region of the mid brain, directly rostral to the pons.
| |
| − | | |
| − | ====Hindbrain====
| |
| − | [[File:Brain sagittal section stem highlighted.svg|thumb|right|300px|Brain sagittal section stem highlighted]]
| |
| − | The hind brain is also called the '''[[CNS Development - Anatomy & Physiology#Development of the Brain|rhombencephalon]]''', it provides the connection between the spinal cord and the rest of the brain. The hindbrain contains many vital structures including the '''Medulla Oblongata''', the '''Pons''' (the link between the cerebellum, forebrain and mid-brain) and the majority of the [[Cranial Nerves - Anatomy & Physiology|'''cranial nerves''']] III to XII. In general the brain stem governs essential functions that are carried out sub-consciously via reflexes. As well as containing numerous cranial nerves, the hind brain also contains many ''''extra-pyramidal pathways'''' which include the '''reticular formation''', the '''olivary nucleus''' and the '''pontine nuclei'''. Nuclei within the hindbrain are also responsible for the '''reflexive control of posture and eye movement'''.
| |
| − | | |
| − | The '''reticular formation''' is a diffuse interconnection of neurons running throughout the brainstem receiving both sensory and motor nerve tracts. This information is then passed on to higher centres in the brain such as the [[cerebrum]]. One important aspect of the reticular formation is that in order to transition from sleep to consciousness, the reticular formation is required to activate the cerebral cortex (ascending reticular activating system). It also contains '''cerebellar pathways''' and peduncles facilitating a connection from the brain stem to the cerebellum. There are also a number of ''''pyramidal pathways'''' and afferent pathways including the '''cuneate''' and '''gracile''' pathways.
| |
| − | =====Medulla Oblongata=====
| |
| − | The medulla oblongata can be found within the '''myelencephalon''' region of the hindbrain. Nuclei in the medulla oblongata control '''heart activity''' including rate and contractility. The medulla oblongata also controls other related functions, including '''blood pressure''' and distribution of blood to different organs. In conjunction with the nuclei found in the pons, the medulla oblongata also exerts an influence on '''respiratory movements'''.
| |
| − | =====Pons=====
| |
| − | The pons can be found in the '''metencephalon''' region of the hindbrain. The pons is able to exert some influence on '''respiratory movements''' and can also influence many '''digestive processes'''.
| |
| − | =====Cranial Nerves=====
| |
| − | [[Cranial Nerves - Anatomy & Physiology|Cranial nerves]] III to XII exit from the brain stem and act to innervate parts of the head, neck, viscera and the thoracic and abdominal cavities. Although most of these nerves contain both sensory and motor fibres, the sensory fibres all have their cell bodies in ganglia outside the brainstem.
| |
| − | =====Cerebellum=====
| |
| − | [[Image:braincerebellumarrow.jpg|thumb|right|150px|The location of the cerebellum. Image courtesy of BioMed Archive]]
| |
| − | The cerebellum is located in the caudal part of the cranial cavity and is caudal to the [[Meninges - Anatomy & Physiology#Dura_mater|'''tentorium cerebelli''']] but dorsal to the fourth ventricle. Its gross structure is made up of a central ''''vermis'''' surrounded by two '''lateral hemispheres'''. It is attached to the brain stem via three pairs of '''peduncles'''; a rostral pair to the midbrain, a middle pair to the pons and a caudal pair to the medulla oblongata. Its internal structure is composed of a cerebellar cortex which is made up of fissures on the surface that divide into lobules and then further sub divide into '''folia''' or leaves. There are white matter fibres running to and from this cortex, also called '''arbor vitae'''. Within the cerebellum there are various '''nuclei''' including the '''dentate, interpositus''' and the '''fastigial'''.
| |
| − | | |
| − | The generalised function of the cerebellum is to receive information regarding any '''movement''' in progress or any intended movement via inputs from the muscles, vestibular system and motor centres of the pyramidal and extrapyramidal systems. The most important function of the cerebellum is to minimise the difference between the intended and the actual movements. The cerebellum then projects corrections regarding these movements to all motor centres of the brain via feedback circuits between the pyramidal and extrapyramidal systems. It should be noted that the cerebellum '''cannot initiate movement'''.
| |
| − | | |
| − | ===Cranial Nerves===
| |
| − | Cranial nerves arise from the brain and [[Hindbrain - Anatomy & Physiology|brain stem]], rather than the spinal cord. Nerves arising from the spinal cord are the [[PNS Structure - Anatomy & Physiology|peripheral nerves]]. There are 12 pairs of cranial nerves and these pairs of nerves passage through [[Skull and Facial Muscles - Anatomy & Physiology|foramina in the skull]], either individually or in groups. Cranial nerves are traditionally referred to by Roman numerals and these numerals begin cranially and run caudally.
| |
| − | The most cranial nerve is the '''Olfactory nerve (I)''' which runs from the nasal cavity through to the olfactory bulb. The next most cranial is the '''Optic nerve (II)''' which runs from the eyes to the [[Forebrain - Anatomy & Physiology#Thalamus|thalamus]]. Cranial nerves III to XII all exit from the brain stem and innervate the head, neck and organs in the thorax and abdomen. In order of most cranial to caudal, these include the '''Oculomotor nerve (III)''', the '''Trochlear nerve (IV)''', the '''Trigeminal nerve (V)''', the '''Abducens nerve (VI)''', the '''Facial nerve (VII)''', the '''Vestibulocochlear nerve (VIII)''', the '''Glossopharyngeal nerve (IX)''', the '''Vagus nerve (X)''', the '''Accessory nerve (XI)''' and the '''Hypoglossal nerve (XII)'''.
| |
| − | | |
| − | Many of the cranial nerves with nuclei within the brain stem contain sensory and motor neurone components. The sensory fibre components have their cell bodies located in ganglia outside the central nervous system and the motor fibre element have their cell bodies within the central nervous system. The'''Olfactory nerve (I)''', '''Optic nerve (II)''' and '''Vestibulocochlear nerve (VIII)''' are sensory nerves. The , '''Oculomotor nerve (III)''', '''Trochlear nerve (IV)''','''Abducens nerve (VI)''','''Accessory nerve (XI)''' and '''Hypoglossal nerve (XII)''' are motor nerves. Finally, the '''Trigeminal nerve (V)''', '''Facial nerve (VII)''','''Glossopharyngeal nerve (IX)''', and '''Vagus nerve (X)''' are mixed sensory and motor nerves.
| |
| − | | |
| − | ====Olfactory Nerve (I)====
| |
| − | The olfactory nerve is involved in the conscious perception of smell. Primary afferent cell bodies are located within the olfactory epithelium of the nasal mucosa on ethmoturbiate bones,rather than in a ganglion like the other cranial nerves. Projections from these cell bodies are the olfactory nerve fibres. The olfactory nerve is a sensory nerve and is composed of many '''Special Visceral Afferent''' fibres. The fibres are formed into bundles that are referred to as 'Olfactory filaments'. The olfactory nerve passes through the [[Skull and Facial Muscles - Anatomy & Physiology#Ethmoid Bone (os ethmoidale)|'''Cribiform plate''']] and is surrounded by meningeal sheets including the [[Meninges - Anatomy & Physiology#Subarachnoid_Space|sub-arachnoid space]]. The olfactory nerve terminates at the [[Forebrain - Anatomy & Physiology#Olfactory_Bulb|olfactory bulb]]. The horse also has nerves which arise from the nasal septum that course into the olfactory bulb, along with the '''vomeronasal nerve''' arising from the '''vomeronasal organ'''. Secondary neurons within the olfactory bulb project through the olfactory tracts to synapse with third order neurons in the medial forebrain bundle, amygdala, septal nuclei and habenular nuclei.
| |
| − | | |
| − | In the horse, special consideration must be given to diseases of the guttural pouch when considering cranial nerve dysfunction. The Glossopharangeal (IX), Vagus (X)and Accessory (XII) nerves are located in the medial compartment of the guttural pouch. The Facial (VII) nerve runs along the lateral compartment. The Mandibular nerve (V2) has limited contact with the dorsal wall of the lateral compartment.. Guttural pouch mycosis commonly results in paresis of cranial nerves IX,V and XII as well as erosion of the internal carotid artery. Rarely, there is involvement of cranial nerves VII and VIII.
| |
| − | | |
| − | ====Optic Nerve (II)====
| |
| − | The optic nerve represents the connection between the receptor cells of the [[Eye - Anatomy & Physiology#The Wall (retina, uvea and sclera)|retina]] and the [[Forebrain - Anatomy & Physiology|forebrain]]. It is not a true nerve, but represents an extension of the brain. The optic nerve is sesory, and is composed of '''Special Somatic Afferent fibres'''.
| |
| − | | |
| − | The '''visual pathway'''' involves three consecutive neurons:
| |
| − | *The first order neuron is the bipolar cells of the [[Eye - Anatomy & Physiology#The Wall (retina, uvea and sclera)|retina]], which are known as rods and cones.
| |
| − | *The second order neuron is the ganglion cells of the retina and axons within the optic nerve. The optic nerve passes through the [[Skull and Facial Muscles - Anatomy & Physiology#Sphenoid Bone (os_sphenoidale)|'''optic chiasm''']], which is an area of the ventral brain where both optic nerves run in a medial direction and eventually decussate (cross). In the horse, approximately 85-88% of fibres decussate. The optic nerve then runs through the [[Skull and Facial Muscles - Anatomy & Physiology#Sphenoid Bone (os_sphenoidale)|'''optic canal''']].
| |
| − | *The third order neuron has its cell body in the lateral geniculate nucleus in the diencephalon. Its axon projects to the visual cortex, which is mostly the contralateral occipital cortex, in the '''optic radiation'''. The occipital lobe is where visual processing takes place at a conscious level.
| |
| − | | |
| − | The nerve is also involved in modulation of '''parasympathetic tone to the iris'''. The first and second order neuron pathways are the same as those responsible for vision, however after synapsing with the lateral geniculate nucleus axons involved in modulation of parasympathetic tone synapse with a third order neuron in the '''pretectal nucleus'''. Most axons from the pretectal nucleus then decussate back to synapse in the parasympathetic component of the '''Occulomotor nerve (III)''' in the ipsilateral eye (because it has crossed once at the optic chiasm and then again at the pretectal nucleus).
| |
| − | | |
| − | The optic nerve can be examined clinically via the [[Neurological Eye Examination - Horse#Menace Response|menace response]] and [[Neurological_Eye_Examination_-_Horse#The_pupillary_light_reflex_(PLR)|pupillary light reflex (PLR)]]. Anopsia (loss of vision) can be seen, especially associated with shear injury to the nerve after head trauma.
| |
| − | | |
| − | ====Oculomotor nerve (III)====
| |
| − | The oculomotor nerve is part of the group of cranial nerves responsible for innervating the [[Skull and Facial Muscles - Anatomy & Physiology#Facial_Muscles|muscles of the head]]. The nerve originates from the ventral [[Midbrain - Anatomy & Physiology|midbrain]] and is a motor nerve. It is composed of '''general somatic efferent fibres''' and '''general visceral efferent fibres'''. The general somatic efferent fibres of the oculomotor nerve are responsible for the motor function of four of the six [[Eye - Anatomy & Physiology#Around_the_Eye|external muscles of the eyeball]]; the 'dorsal rectus', 'medial rectus', 'ventral rectus', 'ventral oblique' and 'levator palpebri superioris' (levator of the upper eyelid). The general visceral efferent fibres of the oculomotor nerve are responsible for the control of pupil diameter and therefore control the 'spincter pupillae' muscle and the 'ciliaris' muscle. These fibres control pupillary constriction via the parasympathetic component of the nerve.
| |
| − | | |
| − | The oculomotor nerve has a pre-ganglionic nucleus in the midbrain and the nerve passes through the [[Skull_and_Facial_Muscles_-_Anatomy_%26_Physiology#Major_Foramen_and_Canals|'''orbital fissure''']], along with the trochlear, abducens and opthalmic branch (V1) of the trigeminal nerve. It synapses in the ciliary ganglion of the eye.
| |
| − | | |
| − | During a clinical examination, horizontal eye movements (strabismus) or an absent [[Neurological_Eye_Examination_-_Horse#The_pupillary_light_reflex_(PLR)|pupillary light reflex (PLR)]] may indicate a problem.
| |
| − | | |
| − | ====Trochlear nerve (IV)====
| |
| − | The trochlear nerve is part of the cranial nerve group responsible for innervation of the [[Skull and Facial Muscles - Anatomy & Physiology#Facial_Muscles|muscles of the head]]. The trochlear nerve originates from the dorsal midbrain and is a motor nerve. It is composed of '''general somatic efferent fibres''' and is the smallest of the cranial nerves.
| |
| − | | |
| − | After leaving the dorsal midbrain, its axons decussate (cross) and then run in a rostral direction through the cavernous sinus before exiting the skill via the [[Skull and Facial Muscles - Anatomy & Physiology#Major Foramen and Canals|'''orbital fissure''']]. In the horse, it may also exit via a seperate trochlear foramen. Finally, it runs to innervate the 'dorsal oblique muscle' muscle of the contralateral eye.
| |
| − | | |
| − | During a clinical examination, a dorso-lateral strabismus may indicate a problem with this nerve.
| |
| − | | |
| − | ====Trigeminal nerve (V)====
| |
| − | The trigeminal nerve is part of the cranial nerve group responsible for innervation of structures originating from branchial arches. The trigeminal nerve nuclei is in the area of the '''pons''' and '''medulla oblongata''' and is the nerve of the 1st branchial arch. The trigeminal nerve provides sensory innervation of cutaneous elements of the face, cornea, mucosa of the nasal septum and mucosa of the oral cavity. It also provides motor fibres to structures also associated with the 1st branchial arch, which are the muscles of mastication (''temporalis'', ''masseter'', ''medial and lateral pterygoids'' and ''rostral digastricus''. There are three primary branches of the trigeminal nerve; the '''Opthalmic nerve (V1)''', the ''' Maxillary nerve (V2)''' and the '''Mandibular nerve (V3)'''.
| |
| − | =====Opthalmic nerve (V1)=====
| |
| − | The opthalmic nerve is a sensory nerve composed of '''general somatic afferent fibres'''. It passes along the cavernous sinus and exits via the [[Skull and Facial Muscles - Anatomy & Physiology#Major Foramen and Canals|'''orbital fissue''']]. As it enters the orbit of the eye, it splits further into the '''lacrimal nerve''', the '''frontal nerve''', the '''nasociliary nerve''' and the '''infratrochlear nerve'''.
| |
| − | * The '''lacrimal nerve''' containes postganglionic parasympathetic fibres from the pterygopalatine ganglion that innervate the lacrimal gland. The lacrimal nerve also contains general somatic afferents that provide sensation to the lateral part of the upper eyelid.
| |
| − | *In the horse, the '''frontal nerve''' exits the medial aspect of the orbit via the '''supraorbital foramen''', becoming the '''supraorbital nerve''', and innervates the upper eyelid and forehead.
| |
| − | *The '''infratrochlear nerve''' innervates the medial aspects of the eyelids, third eyelid and frontal sinus.
| |
| − | * '''Nasociliary nerves''', which carry parasympathetic fibres from the oculomotor nerve to the iris, also provide sensory innervation to the globe.
| |
| − | | |
| − | =====Maxillary nerve (V2)=====
| |
| − | The maxillary nerve is a sensory nerve composed of '''general somatic afferent fibres'''. The maxillary nerve passes along the cavernous sinus and exits through the [[Skull and Facial Muscles - Anatomy & Physiology#Major Foramen and Canals|'''round foramen''']] before entering the [[Skull and Facial Muscles - Anatomy & Physiology#Major Foramen and Canals|'''alar canal''']]. It also runs across the wall of the '''pterygopalatine fossa''' and enters the '''infraorbital canal''' via the '''maxillary foramen'''. Whilst in the infraorbital canal, the maxillary nerve branch then branches further into the '''infraorbital nerve''' which supplies sensory fibres to the upper dental arcade. On exiting the infraorbital canal via the infraorbital foramen, the maxillary nerve branches again into the '''zygomatic nerve''' and '''pterygopalatine nerve''' supplying sensory fibres to the palate, lower eyelid, upper lip, nasal planum, and dorsal face.
| |
| − | | |
| − | =====Mandibular nerve (V3)=====
| |
| − | The mandibular nerve is a mixed sensory '''general somatic afferent fibres''' and motor '''general somatic efferent''' nerves. The mandibular nerve passes through the '''foramen lacerum''' in the horse. It provides motor branches to the [[Mastication|masticatory muscles]], the [[Larynx - Anatomy & Physiology#Intrinsic Musculature|ventral throat]] and [[Tongue - Anatomy & Physiology#Muscles|muscles of the palate]]. The mandibular nerve further branches into the '''masticatory nerve''', '''masseteric nerve''' and the '''temporal nerve'''. The mandibular nerve provides sensory branches called the '''buccal nerve''', '''auriculotemporal nerve''', and then itself divides into two smaller branches; the '''lingual nerve''' and the '''inferior alveolar nerve'''. The auriculotemporal nerve carries sensory information from the middle ear, temporal area and portions of the guttural pouch. The '''lingual nerve''' receives sensory taste fibres and also connects some sensory taste fibres to parasympathetic salivary glands via the [[Tongue - Anatomy & Physiology#Innervation|'''chorda tympani''']]. Via the chorda tympani branch, the mandibular branch supplies sensory fibres related to taste to the rostral 2/3 of the tongue. The lingual branch of the glossopharyngeal nerve supplies sensory fibres to the caudal 1/3 of the tongue.
| |
| − | | |
| − | ====Abducent nerve (VI)====
| |
| − | The abducent nerve is part of the cranial nerve group responsible for innervation of the [[Skull and Facial Muscles - Anatomy & Physiology#Facial_Muscles|muscles of the head]]. The abducent nerve originates from the medulla oblongata and is a motor nerve. It is composed of '''general somatic efferent fibres''' which are responsible for controlling the ''lateral rectus'' and ''retractor bulbi'' muscles of the eye. The nerve passes through the '''orbital fissure''' and can be found within the same layer of the meninges as the opthalmic branch (V1) of the trigeminal nerve (V).
| |
| | | | |
| − | During a clinical examination, medial strabismus may indicate a problem with this nerve.
| + | A single Schwann cell forms a single myelin sheath or internode and there is a reasonably constant relationship between the myelin thickness and the internodal length, which in turn is associated with axon calibre. Large axons have long, thick myelin sheaths and therefore conduct more rapidly. The internodes do not abut one another but are separated by an exposed area of axon called the '''node of Ranvier'''. If the axons remain of small diameter, then a Schwann cell will continue to associate with many axons, although none of them are myelinated. Thus, ''even unmyelinated axons retain a Schwann cell ensheathment''. These non-myelinating Schwann cells are sometimes referred to as ''Remak cells.'' |
| | + | ===Axoplasmic Transport=== |
| | + | Neurons are very large cells and most of a neurons cytoplasm is present in its processes while most of the cells RNA is located in cell body (Nissl substance). These cells have therefore evolved mechanisms to transport large macromolecules and organelles up and down processes. |
| | + | ===Anterograde Transport=== |
| | + | Anterograde transport moves substances from the cell body to the axon. Two basic forms of anterograde transport can be recognised: '''fast anterograde transport''' and '''slow anterograde transport'''. Fast anterograde transport allows movement of all membranous organelles such as synaptic vesicles and occurs at a rate of around 400mm/day (recent evidence suggests that there are many forms of fast anterograde transport, mediated by different kinesins). Fast anterograde transport depends critically on oxidative metabolism, and is, in fact independent of the cell body. Microtubules act as a static track along which the organelles can move, driven by the ATPase '''kinesin''' which acts as a "motor" molecule. Fast anterograde transport is independent of the cell body. Anything which interferes with energy supply or cytoskeleton necessary for fast anterograde transport has profound effects on the health of the axon. Agents such as colchicine or vincristine block microtubule assembly, disrupting fast anterograde transport. '''Slow anterograde transport''' deals with cytoskeletal elements and large soluble proteins. Slow anterograde transport can be further sub-divided into a slow component, which occurs at about 2mm/day (neurofilament, rubulin, actin) and a fast component, which occurs at around 4 mm/day, transporting all other proteins (eg myosin, clathrin). |
| | + | ===Retrograde Transport=== |
| | + | Retrograde transport returns materials from the axon terminal to the cell body, either for degradation or restoration and reuse. As with fast anterograde transport, particles move along microtubules. The motor molecule for retrograde transport is '''dynein''' which is a microtubule-associated ATPase. The retrograde transport system is important not only for returning material to the cell body, but also provides the means whereby target-derived trophic factors, such as nerve growth factor (NGF) for dorsal root ganglion neurons, are conveyed to the cell body where they promote cell survival. Research is being undertaken into the use of trophic factors to promote cell survival during degenerative pathology. The retrograde transport system can be "hijacked" by harmful substances to gain entry to the peripheral neuron and ultimately the CNS. [[Herpesviridae|Herpes virus]], [[Tetanus - Horse|tetanus]] and heavy metals all affect the retrograde transport system. |
| | + | ==Blood Supply== |
| | + | The epineurium is penetrated by the vascular supply to the nerve and this blood supply is known as the '''vasa nervorum'''. Only capillaries occur within the endoneurial compartment. The capillaries of the endoneurium are joined by tight junctions and provide a barrier to large macromolecules. This forms the basis of the blood-nerve barrier (BNB), which has similarities to the [[Blood Brain Barrier - Anatomy & Physiology|blood-brain barrier]] of the CNS. The BNB appears to be relatively weak in the sensory ganglia because fenestrations occur between endothelial cells in this location. Sensory ganglia are therefore more vulnerable to blood-borne agents. A further "barrier" is provided by the perineurium which consists of sheets of flattened cells, connected by tight junctions and covered on both sides by a basal lamina. The only route across this structure is trans- rather than inter-cellular. |
| | | | |
| − | ====Facial nerve (VII)====
| + | [[Category:To Do - AP Review]] |
| − | The facial nerve is part of the cranial nerve group responsible for the innervation of structures originating from the branchial arches. It originates from the '''medulla oblongata''' and from the second branchial arch. It has a common dura sheet with the opthalmic (V1) branch of the trigeminal nerve. The facial nerve is of a mixed composite, made up of a number of different fibre types. It has a '''general somatic efferent fibre''' within the ear canal, a '''general visceral efferent fibre''' acting under parasympathetic control to some salivary glands, lacrimal glands, nasal cavity and palate, a '''special visceral afferent fibre''' providing taste to the rostral 2/3 of the tongue and finally it has a '''general somatic efferent fibre''' supplying motor function to the muscles of facial expression and ''caudal digastricus''.
| |
| − | | |
| − | The facial nerve enters the petrosal bone via the [[Skull and Facial Muscles - Anatomy & Physiology#Temporal Bone (os_temporale)|'''internal acoustic meatus''']] along with the vestibulocochlear nerve. The facial nerve also runs inside the '''facial canal'''. There are a number of intermediate branches which separate from the main facial nerve inside the facial canal including the '''greater petrosal nerve''', '''stapedial nerve''' (motor) and the '''chorda tympani'''. These then emerge via the [[Skull and Facial Muscles - Anatomy & Physiology#Major Foramen and Canals|'''stylomastoid foramen''']] at the caudoventral aspect of the skull. The chorda tympani of the facial nerve represents the '''special visceral afferent fibre''' supplying taste to the rostral 2/3 of the tongue.
| |
| − | | |
| − | There are also numerous external branches of the facial nerve once the facial nerve has left the facial canal. These include the '''internal auricular nerve''', the '''auriculopalpebral nerve''', the '''rostral auricular nerve''', the '''palpebral nerve''', the '''dorsal buccolabial nerve''', the '''ventral buccolabial nerve''', the '''ramus colli''', the '''digastric nerve''', the '''stylohoid nerve''' and the '''caudal auricular nerve'''.
| |
| − | | |
| − | The facial nerve supplies motor innervation to the muscles of facial expression. These are superficial flat, thin muscles that originate from bony areas of fascia and then radiate out around the skin. They may also often from sphincters such as around the mouth and eye.
| |
| − | | |
| − | During a clinical examination any facial paralysis, drooling or abscence of blinking may indicate a problem with the facial nerve.
| |
| − | | |
| − | ====Vestibulocochlear nerve (VIII)====
| |
| − | The vestibulocochlear nerve is part of the special senses group of cranial nerves and is made up of two components; the vestibular nerve and the cochlear nerve. The vestibular nerve is responsible for balance whilst the cochlear nerve is responsible for hearing. The nerves send impulses from the inner ear which contains the [[Ear - Anatomy & Physiology#Vestibular Receptors and Balance|vestibular apparatus]] and [[Ear - Anatomy & Physiology#The Cochlea|cochlea]]. The vestibulocochlear nerve is a sensory nerve made up of '''special somatic afferent fibres'''. It passes through the '''internal acoustic meatus''' and into the '''petrosal bone'''. The facial nerve also takes this route.
| |
| − | | |
| − | Clinical problems with the vestibulocochlear nerve would be indicated on examination by changes in hearing and/or strabismus and [[Vestibular System Examination|nystagmus]]. A head tilt is also associated with this nerve.
| |
| − | | |
| − | ====Glossopharyngeal nerve (IX)====
| |
| − | The glossopharyngeal nerve is part of the group of cranial nerves responsible for innervation of structures derived from the branchial arches. This nerve innervates structures related to the third branchial arch. It is also part of a group, together with the vagus and accessory nerves, that passes through the '''jugular foramen''' which is termed the '''vagus group'''. The glossopharyngeal nerve has cell bodies that are referred to as '''nucleus ambiguus'''. The glossopharyngeal nerve originates from the '''medulla oblongata''' and has several branches including the '''pharyngeal nerve''', the '''lingual nerve''' and the '''tympanic branches'''.
| |
| − | | |
| − | The glossopharyngeal nerve is composed of many fibre types including '''general somatic efferent fibres''' that innervate the stylopharyngeus muscle; the '''general visceral afferent fibres''' that provide sensory information from the carotid body, the pharynx and the middle ear; the '''general visceral efferent fibres''' that provide parasympathetic innervation to the parotid and zygomatic salivary glands; the '''special visceral afferent fibres''' that provide taste caudal to the tongue and finally the '''general somatic afferent fibres''' that provide sensory information from the external ear. The '''lingual branch''' of the glossopharyngeal nerve provides '''general somatic afferent fibres''' and '''special visceral afferent fibres''' to the caudal 1/3 of the tongue.
| |
| − | | |
| − | On clinical examination, choking or dysphagia as a result of malfunctioning or absent pharyngeal reflexes would indicate a problem with the glossopharyngeal nerve.
| |
| − | | |
| − | ====Vagus nerve (X)====
| |
| − | The vagus nerve is part of the group of cranial nerves responsible for innervation of structures derived from the branchial arches. It is also part of a group, together with the glossopharyngeal and accessory nerves, that passes through the '''jugular foramen''' which is termed the '''vagus group'''. The vagus nerve innervates structures related to the fourth branchial arch. The vagus nerve has cell bodies that are referred to as '''nucleus ambiguus'''.
| |
| − | | |
| − | The vagus nerve is composed of many different types of nerve fibre including '''general somatic efferent fibres''' supplying motor function to the muscles of the larynx, pharynx, palate and oesophagus; '''general visceral afferent fibres''' to the base of the tongue, pharynx and larynx; '''general visceral efferent fibres''' for parasympathetic supply of the thoracic and abdominal viscera; '''special visceral afferent fibres''' supplying taste to regions of the epiglottis and palate and finally '''general somatic afferent fibres''' to the external ear and the dura mater. The vagus nerve also supplies '''general somatic afferent fibres''' and '''special visceral afferent fibres''' to the root of the tongue.
| |
| − | | |
| − | There are many functional components of the vagus nerve including the heart, larynx, pharynx and many other viscera. On clinical examination any changes related to gag reflexes, blood pressure or heart rate, changes in 'voice' (dysphonia) or inspiratory dyspnoea may indicate a problem with the vagus nerve.
| |
| − | | |
| − | ====Accessory nerve (XI)====
| |
| − | The accessory nerve is part of the group of cranial nerves responsible for innervation of structures derived from the branchial arches. It is also part of a group, together with the glossopharyngeal and vagus, nerves that passes through the '''jugular foramen''' which is termed the '''vagus group'''. The accessory nerve supplies structures related to the fourth branchial arch. The accessory nerve has cell bodies that are referred to as '''nucleus ambiguus''' and originate in the '''medulla oblongata'''. The cranial root of the accessory nerve actually contributes to the vagus nerve and to the striated muscles of the pharynx, larynx, palate and oesophagus.
| |
| − |
| |
| − | However, the accessory nerve also contributes to the cervical spinal cord and spinal root through the '''foramen magnum'''; providing innervation to muscles of the neck. The spinal root of the accessory nerve branches into the '''dorsal branch''' and the '''ventral branch'''. The dorsal branch innervates the ''brachiocephalicus'', ''trapezius'' and ''omotransversarius'' muscles of the dorsal neck. The ventral branch innervates the ''sternocephalicus'' muscle.
| |
| − | | |
| − | During clinical examination any difficulties in turning the neck or muscle atrophy around the dorsal and ventral neck may indicate a problem with the accessory nerve.
| |
| − | | |
| − | ====Hypoglossal nerve (XII)====
| |
| − | The hypoglossal nerve is part of the group of cranial nerves responsible for the control of muscles of the head. It is in part a cervical nerve due to its caudal position on the [[Hindbrain - Anatomy & Physiology|brain stem]]. The nerve is composed of '''general somatic efferent fibres''' which control the intrinsic and extrinsic muscles of the tongue (together with other nerves including the lingual nerve, facial nerve, lingual branch of the glossopharyngeal nerve and the vagus nerve). The nucleus of the nerve is located within the ''medulla oblongata'' of the brain stem and it passes through the '''hypoglossal canal'''.
| |
| − | | |
| − | During a clinical examination any deviation of the tongue may indicate a problem with this nerve. Deviation of the tongue is always to the side of the lesion initially.
| |
| − | | |
| − | ===Spinal Cord===
| |
| − | The '''spinal cord''' is constructed of the ''[[Spinal Cord - Anatomy & Physiology#Marginal layer|marginal layer]] ''which has axons and white matter, the ''[[Spinal Cord - Anatomy & Physiology#Mantle|mantle]]'' which contains cell bodies and grey matter and the ''[[Spinal Cord - Anatomy & Physiology#Spinal Canal|spinal canal]]''. This canal conducts sensory information from the peripheral nervous system (both somatic and autonomic) to the brain, conducts motor information from the brain to various effectors and acts as a minor reflex center. The spinal cord can be divided to several regions:
| |
| − | :'''cervical''' ''(C1-C6)''
| |
| − | :'''cervicothoracic''' ''(C7-T2)''
| |
| − | :'''thoracolumbar''' ''(T3-L3)'' | |
| − | :'''lumbosacral''' ''(L3-S2)''
| |
| − | :'''sacral''' ''(S3 onwards)''
| |
| − | | |
| − | Nerves originating from the spinal cord and the segmental spinal nerves innervate the limbs.
| |
| − | <br> <br>
| |
| − | The '''forelimb nerves''' include the '''suprascapular''' ''(C5-6)'', the '''musculocutaneous''' ''(C5-7)'', the '''ulna/median''' (Originates from the brachial plexus, which is formed from ''C5-T1'') and the '''radial''' ''(C5-T1)''.
| |
| − | | |
| − | The '''hindlimb nerves''' include the '''obturator''' ''(L2-4)'', the '''femoral''' ''(L2-4)'' and the '''sciatic''' ''(L4-S3)''. The sciatic nerve branches to the tibial nerve and the peroneal nerve.
| |
| − | <br> <br>
| |
| − | | |
| − | ====Sensory Pathways====
| |
| − | [[File:Spinal cord tracts - English.png|right|200px|thumb|Spinal cord tracts]]
| |
| − | The spinal cord contains a number of [[Sensory Pathways - Anatomy & Physiology|sensory (ascending) pathways]] or tracts contained within the [[Central Nervous System - Anatomy & Physiology#White Matter|white matter]]. These pathways allow sensory information such as pain, touch, temperature or kinaesthesia (conscious proprioception) to be passed through the spinal cord and on to higher levels of the brain.
| |
| − | <br />
| |
| − | | |
| − | ====Vasculature====
| |
| − | It is important to note that there is no direct vasculature to the spinal cord but instead there are a number of choroid plexuses that act as an exchanger between the vasculature of the spinal cord/brain and the fluid surrounding these structures. This distinction is referred to as the [[Blood Brain Barrier - Anatomy & Physiology|"blood-brain barrier"]].
| |
| − | <br />
| |
| − | <br />
| |
| − | The vasculature of the spinal cord has a close relationship with the [[Cerebral Spinal Fluid - Anatomy & Physiology|cerebrospinal fluid (CSF)]] within the subarachnoid space. This CSF effectively forms a water jacket that buoys up the spinal cord and protects it from external influences. Therefore it is extremely important that the CSF has the appropriate properties in order to undertake this role. The vasculature of the spinal cord therefore has to provide the appropriate level of oxygen, pressure, pH and nutrients to maintain homeostasis of the spinal cord. As the CSF also performs this role within the skull, the vasculature of the brain has an important relationship with every aspect of the ventricles and subarachnoid space within the central nervous system.
| |
| − | <br />
| |
| − | | |
| − | =====Arterial Supply=====
| |
| − | The spinal cord is supplied by three main arteries that run along its length; the '''Ventral Spinal Artery''', and paired '''Dorsolateral Spinal Arteries'''. The ventral spinal artery is the largest and follows the ventral fissure of the spinal cord. The dorsolateral arteries run close to the groove from which the dorsal nerve roots arise. Together with these three main arteries, the spinal cord is also supplied by branches from regional arteries including branches in the cervical, intercostal, lumbar and sacral regions. These regional arteries enter the spine at the intervertebral foramina, often accompanying the roots of spinal nerves. These regional arteries also form plexuses into which the three main longitudinal arteries run. The number and type of arteries that enter the spine from regional branches varies with species and also between individuals.
| |
| − | <br />
| |
| − | The '''ventral spinal artery''' supplies the main "core" of the spinal cord, i.e. the [[Central Nervous System - Anatomy & Physiology#Grey Matter|grey matter]]. It also partially supplies the white matter via the ventral fissure, although the majority of the white matter is supplied by radial branches of the dorsolateral arteries. There are also a number of anastamoses between both sets of arteries.
| |
| − | | |
| − | =====Venous Supply=====
| |
| − | Along the length of the spinal cord runs the vertebral venous plexus which drains the blood from the vertebrae and surrounding musculature. This venous plexus gives rise to veins that then leave the vertebrae via the intervertebral foramina and then go on to join the major venous channels of the neck and the trunk; namely the vertebral, cranial caval, azygous and caudal caval veins.
| |
| − | <br />
| |
| − | <br />
| |
| − | The venous plexus consists of paired channels within the epidural space that lie in a '''ventral''' position to the spinal cord. Each side of the pair is connected to its opposing plexus around the vertebrae resulting in a ladder-type pattern of venous vessels. The connections between each side are via the intervertebral foramina and these vessels are in close proximity to the spinal nerves.
| |
| − | <br />
| |
| − | <br />
| |
| − | The veins around the plexuses have no valves and can theoretically pass blood in either direction. The vessels are able to adjust their size/pressure to compensate for intrathoracic pressure. This intermittency of flow causes an increased risk of septic or neoplastic disease within the vertebral column. Where blood is impeded or where flow may become temporarily held stagnant, this may allow tumor seeds or micro-organisms to settle within tributaries.
| |
| − | | |
| − | ====Lymphatics====
| |
| − | There are no lymphatic vessels or nodes within the spinal cord or other central nervous tissue.
| |
| − | | |
| − | ===Meninges===
| |
| − | The meninges are layers of tissue surrounding the central nervous system (CNS). Meningitis is the inflammation of these layers. Gaps and spaces between the meninges are named.
| |
| − | ====Dura mater====
| |
| − | The Dura mater is the outer most layer and is made up of a dense fibrous connective tissue. The space in the vertebral canal ouside the dura mater is the '''epidural space'''. In the cranium, the dura layer is fused with the periosteum and therefore is in effect single layer without an epidural space. The dura contains a number of folds throughout its coverage of the brain including the '''falx cerebri''', a midline fold between cerebral hemispheres, the '''tentorium cerebelli''', an oblique fold between the cerebrum and cerebellum and the '''diaphragma sellae''' which forms a collar around the neck of the pituitary and forms the roof of the hypophyseal fossa. This layer and these associated folds all provide structural support to the brain and prevent the brain from undergoing excess movement within the skull. Where the dura mater folds between brain tissues it splits into two distinct layers that are separated by large blood filled spaces called '''venous sinuses'''. Venous sinuses are directly connected to the venous system and venous blood from vessels supplying the brain return to the heart via these sinuses.
| |
| − | | |
| − | ====Subdural space====
| |
| − | The subdural space lies between the dura mater and the next meningial layer, the arachnoid mater. The subdural space is narrow potential space, where the two meningeal leayers lie in close proximity; but do not meet. The subdural space is thought to contain only lymph-like fluid. The meningeal layers can move apart in the event of injury or increased pressure; for example pooling of blood in the subdural space (subdural haematoma).
| |
| − | ====Arachnoid mater====
| |
| − | This is the middle meningial layer and lies between the dura mater and the pia mater, the innermost meningeal layer. The arachnoid mater is a delicate structure and is constructed with non-vascular connective tissue. This layer also has small protrusions through the dura mater into the previously mentioned venous sinuses called '''Arachnoid villus''' and these allow [[Cerebral_Spinal_Fluid_-_Anatomy_&_Physiology|cerebrospinal fluid]] (CSF) to enter and exit the blood stream. These protrusions adhere to the inner surface of the skull via ''calvaria'' processes.
| |
| − | ====Subarachnoid Space====
| |
| − | The subarachnoid space lies between the arachnoid mater and pia mater. Both meninges are connected via a fine network of connective tissue filaments (spider web-like) which run through the space, originating from the arachnoid mater. This space also contains '''cerebrospinal fluid (CSF)''' from ventricular system. The largest parts of this space are called the ''cisterns'', which are used for the collection of CSF. For example there is a '''cerebellomedullary cistern''' around the foramen magnum.
| |
| − | ====Pia Mater====
| |
| − | This is the innermost layer and is firmly bound to the underlying neural tissue of the brain and spinal cord. The inner surface of the brain facing this meningial layer is lined with ependymal cells. The pia mater is highly vascular and is formed from delicate connective tissue. It also contains arteries and veins, but not venous sinuses.
| |
| − | ===Cerebrospinal Fluid===
| |
| − | Cerebrospinal fluid ('''CSF''') surrounds the brain and spinal cord. It helps cushion the central nervous system (CNS), acting in a similar manner to a shock absorber. It also acts as a chemical buffer providing immunological protection and a transport system for waste products and nutrients. The CSF also provides buoyancy to the soft neural tissues which effectively allows the neural tissue to "float" in the CSF. This prevents the brain tissue from becoming deformed under its own weight. It acts as a diffusion medium for the transport of neurotransmitters and neuroendocrine substances.
| |
| − | ====Production====
| |
| − | CSF is a clear fluid produced by dialysis of blood in the '''choroid plexus'''. Choroid plexi are found in each lateral ventricle and a pair are in the third and fourth ventricle. Further production also comes from the '''ependymal cell''' linings and vessels within the pia mater.
| |
| − | | |
| − | Edendymal cell production of CSF is via ultrafiltration of blood plasma and active transport across the ependymal cells. The ependyma is connected via a series of tight junctions preventing molecules passing between cells. The ependyma also sits on a basement membrane to provide support to the ependymal cells and provide further protection against blood perfusion. In areas of the brain where there are choroid plexi, the endothelium of the plexus vessel sits immediately adjacent to the basement membrane of the ependymal cells. Of the total CSF production, 35% is produced within the third ventricle of the brain, 23% via the fourth ventricle and 42% from general ependymal cell filtration.
| |
| − | | |
| − | CSF has a very low protein constituent, with only albumin being present together with a very low level of cellularity. The biochemistry of CSF includes high concentrations of sodium and chloride and very high concentrations of magnesium. Concentrations of potassium, calcium and glucose are low.
| |
| − | | |
| − | ====Circulation====
| |
| − | Once produced, CSF is then circulated, due to hydrostatic pressure, from the choroid plexus of the '''lateral ventricles''', through the '''interventricular foramina''' into the '''3rd ventricle'''. The lateral ventricles are paired and are located in the cerebral hemispheres. The 3rd ventricle is located in the diencephalon and surrounds the thalamus. CSF then flows through the '''cerebral aqueduct''' (aqueduct of Sylvius or mesencephalic aqueduct) into the '''4th ventricle'''. The 4th ventricle is located in the hindbrain. From the 4th ventricle the CSF may flow down the central canal of the spinal cord, or circulate in the '''subarachnoid space'''. The central canal of the spinal cord is in direct communication with the 4th ventricle. Most CSF escapes from the ventricular system at the hindbrain '''Foramen of Luschka''' (lateral apertures) into the subarachnoid space. Once in the subarachnoid space, the CSF may enter the '''cerebromedullary cistern''' (a dilation of the subarachnoid space between the cerebellum and the medulla) and then circulate over the cerebral hemispheres. CSF also flows down the length of the spinal cord in the subarachnoid space. Another dilation of the subarachnoid space occurs caudally due to the dura and arachnoid meninges continuing on past the end of the spinal cord. This gives rise to the '''lumbar cistern'''.
| |
| − | | |
| − | Large amounts of CSF are drained into venous sinuses through arachnoid granulations in the '''dorsal sagittal sinus'''. The dorsal sagittal sinus is located between the folds of dura, known as the '''falx cerebri''', covering each of the cerebral hemispheres. Arachnoid granulations contain many villi that are able to act as a one way valve helping to regulate pressure within the CSF, and these '''arachnoid villi''' push through the dura and into the venous sinuses.
| |
| − | ===Vasculature===
| |
| − | | |
| − | ==Peripheral Nervous System==
| |
Central Nervous System
Clinical Links
Peripheral Nervous System
Introduction
The Peripheral Nervous System is made up of cranial and spinal nerves. Spinal nerves are named after the vertebra immediately above it, except for cervical vertebra. There are 7 cervical vertebrae and 8 cervical spinal nerves. The peripheral nervous system can be divided into the somatic nervous system and autonomic nervous system. The somatic nervous system co-ordinates body movements and also receives external stimuli. It regulates activities that are under conscious control. The autonomic nervous system subdivided into the sympathetic nervous system, parasympathetic nervous system, and enteric division. The sympathetic nervous system is the ‘fight or flight’ system which comes into role when an animal is under threat, its main neurotransmitter is adrenaline. The parasympathetic nervous system is the ‘rest and digest’ system which is responsible for digestion. Its main neurotransmitter is acetylcholine.
Structure
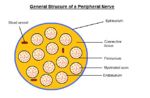
Peripheral Nerve Structure. Sophie Stenner, RVC 2008
Nerve fibres reside in a connective tissue matrix called the endoneurium and are gathered together into bundles or fascicles defined by a second connective tissue layer called the perineurium. Groups of fascicles are then gathered together in a third connective tissue layer called the epineurium. Thus, peripheral nerves have a three-tiered hierarchical arrangement of connective tissue. Renaut bodies are loose, cigar-shaped whorls of extracellular matrix within fascicles that are common in equine nerves at points of stress or compression.
Nerve Fibre
The nerve fibre consists of the impulse-carrying axon, which is surrounded by an ensheathing cell, the Schwann cell, which in turn is surrounded by an acellular basal lamina that is continuous along the length of the nerve. Nerve fibres come in various discrete diameter groups, which are reflected in their conduction velocities. The larger the diameter the more rapid the rate of impulse conduction. Particular targets or receptors are associated with axons of a particular diameter. Axons connected to muscles spindles have a large diameter (20 µm) and conduct at 120 m/s whilst the smallest myelinated fibres are about 1µm and conduct at around 6 m/s. The smallest fibres of all are the unmyelinated fibres (the high-threshold sensory afferents, or C-fibres, and post-ganglionic autonomies) and have a diameter of between 1 and 0.1 µm. These fibres do not conduct by saltatory conduction and have very slow conduction rates of around 0.5 m/s.
Axon
Axons have an outer membrane called the axolemma and within this there is the axoplasm which is continuous with the cytoplasm of the neuron. There are no ribosomes, either free or attached to endoplasmic reticulum in axons and therefore, no protein synthesis. Protein synthesis takes place within the cell body and some dendrites and all protein replacement required for the maintenance of the axon depends on proteins being imported from the cell body. A critical feature of the axon is its cytoskeleton, which consists of two key elements; neurofilaments and microtubules. Neurofilaments are intermediate filaments of about 10 nm diameter, and belong to the same class as other cytoskeletal proteins such as keratin, desmin, vimentin, or GFAP of astrocytes. Neurofilaments are formed from a triplet of polypeptide subunits of heavy (~ 200 kD), medium (~ 150 kD) and low (~ 60 kD) molecular weights. Typically, these subunits are heavily phosphorylated and are more numerous than microtubules, especially in large diameter axons, having a pivotal role in determining axon diameter. They are formed in the cell body, transported down the axon by axoplasmic transport and degraded in the terminals by Ca2+ activated proteases. In other words, there is a constant turnover of neurofilament within the healthy axon. Microtubules within axons are similar to microtubules elsewhere, consisting of polymerised dimers of alpha and beta tubulin arranged as a hollow tube of about 28 nm. They are relatively abundant in smaller diameter axons, and are also synthesised in the cell body. An important component of the cytoskeleton are the microtubule associated proteins or MAP's and the tau proteins. These proteins are important in microtubule assembly and stability. Different classes of MAP's occur in the dendrites and the axons, and to some extent account for the different ultrastructural features that distinguish these two types of neuronal process. They form cross links between adjacent microtubules but also connect to neurofilaments and actin microfilaments, implying complex interactions between the various components of the axon skeleton.
Schwann Cell
Myelination in the PNS is achieved by the Schwann cell, a derivative of neural crest cells, which bud off from the neuroepithelium at a very early stage of neurogenesis. During development, Schwann cells engage many small axons and as axonal diameter increases, Schwann cells eventually relate with only a single axon c.f oligodendrocytes. This single axon is enveloped in a trough by the Schwann cell processes that engulf it and as the processes come together, an inner mesaxon is formed. The leading-edge process continues to move over the axon forming a spiral. Myelination, an extremely complex molecular process, occurs when the cytoplasm within the process is extruded allowing the internal surfaces of the membrane to come together as the major dense line, the outer membrane apposition constituting the intraperiod line. The alternating pattern of these two form the lamellae of compacted myelin. The myelin sheath is attached to, and is an integral part of the Schwann cell on which it is dependent for its maintenance.
A single Schwann cell forms a single myelin sheath or internode and there is a reasonably constant relationship between the myelin thickness and the internodal length, which in turn is associated with axon calibre. Large axons have long, thick myelin sheaths and therefore conduct more rapidly. The internodes do not abut one another but are separated by an exposed area of axon called the node of Ranvier. If the axons remain of small diameter, then a Schwann cell will continue to associate with many axons, although none of them are myelinated. Thus, even unmyelinated axons retain a Schwann cell ensheathment. These non-myelinating Schwann cells are sometimes referred to as Remak cells.
Axoplasmic Transport
Neurons are very large cells and most of a neurons cytoplasm is present in its processes while most of the cells RNA is located in cell body (Nissl substance). These cells have therefore evolved mechanisms to transport large macromolecules and organelles up and down processes.
Anterograde Transport
Anterograde transport moves substances from the cell body to the axon. Two basic forms of anterograde transport can be recognised: fast anterograde transport and slow anterograde transport. Fast anterograde transport allows movement of all membranous organelles such as synaptic vesicles and occurs at a rate of around 400mm/day (recent evidence suggests that there are many forms of fast anterograde transport, mediated by different kinesins). Fast anterograde transport depends critically on oxidative metabolism, and is, in fact independent of the cell body. Microtubules act as a static track along which the organelles can move, driven by the ATPase kinesin which acts as a "motor" molecule. Fast anterograde transport is independent of the cell body. Anything which interferes with energy supply or cytoskeleton necessary for fast anterograde transport has profound effects on the health of the axon. Agents such as colchicine or vincristine block microtubule assembly, disrupting fast anterograde transport. Slow anterograde transport deals with cytoskeletal elements and large soluble proteins. Slow anterograde transport can be further sub-divided into a slow component, which occurs at about 2mm/day (neurofilament, rubulin, actin) and a fast component, which occurs at around 4 mm/day, transporting all other proteins (eg myosin, clathrin).
Retrograde Transport
Retrograde transport returns materials from the axon terminal to the cell body, either for degradation or restoration and reuse. As with fast anterograde transport, particles move along microtubules. The motor molecule for retrograde transport is dynein which is a microtubule-associated ATPase. The retrograde transport system is important not only for returning material to the cell body, but also provides the means whereby target-derived trophic factors, such as nerve growth factor (NGF) for dorsal root ganglion neurons, are conveyed to the cell body where they promote cell survival. Research is being undertaken into the use of trophic factors to promote cell survival during degenerative pathology. The retrograde transport system can be "hijacked" by harmful substances to gain entry to the peripheral neuron and ultimately the CNS. Herpes virus, tetanus and heavy metals all affect the retrograde transport system.
Blood Supply
The epineurium is penetrated by the vascular supply to the nerve and this blood supply is known as the vasa nervorum. Only capillaries occur within the endoneurial compartment. The capillaries of the endoneurium are joined by tight junctions and provide a barrier to large macromolecules. This forms the basis of the blood-nerve barrier (BNB), which has similarities to the blood-brain barrier of the CNS. The BNB appears to be relatively weak in the sensory ganglia because fenestrations occur between endothelial cells in this location. Sensory ganglia are therefore more vulnerable to blood-borne agents. A further "barrier" is provided by the perineurium which consists of sheets of flattened cells, connected by tight junctions and covered on both sides by a basal lamina. The only route across this structure is trans- rather than inter-cellular.
